Races, Human. All human beings are descended from people who lived hundreds of thousands of years ago. Thus, we all share a common ancestry. This means that all people living today are related to one another. But even though we are all related, we do not all look alike. Our bodies have different sizes and shapes, our skins have varying shades, our eyes differ in color and shape, our lips and noses have different shapes, and our hair has different colors and textures.
Most anthropologists believe that human beings originated in Africa and gradually spread throughout the world (see Prehistoric people (Migration from Africa)). They have observed that groups of people who have lived in certain parts of the world for many thousands of years tend to differ from groups living in other parts of the world. Living in regions with differing environments is one reason human beings have developed different appearances. For example, people whose ancestors lived for many generations in northern parts of the world—such as northern Europe or northern Japan—tend to have light-colored skin. People who come from places near the equator, such as central Africa or southern India, tend to have dark-colored skin. People who come from places between those two environmental extremes tend to have medium-colored skin. For information on how skin colors result from adaptations to the environment, see the Climatic adaptations section of this article.
In some instances, we observe that certain physical traits tend to cluster in a group. For example, we might associate blond hair, blue eyes, and fair skin with people from Denmark, Norway, and Sweden. We also might associate red hair, green eyes, and a freckled complexion with people from Ireland. However, many people in these four countries actually have brown hair, brown eyes, and light brown skin. This example shows some of the problems facing human biologists who attempt to classify human beings into races.
Biologists define a race as a subdivision of a plant or animal species. The members of the same species resemble one another in many essential ways. Most importantly, they can breed with one another and produce fertile offspring. Members of different species usually cannot interbreed and produce fertile offspring. Grizzly bears and black bears, for example, are closely related North American bears. Despite their similarities, grizzly bears and black bears do not interbreed. Therefore, they belong to different species.
Many plant and animal species can be subdivided into groups that differ from one another. These groups are often called subspecies. Among grizzly bears, for instance, biologists observe distinct physical differences from region to region. They group grizzly bears into subspecies based on these differences.
All living human beings belong to the subspecies Homo sapiens sapiens. But like those of the grizzly bear, human populations differ from one region to another. Scholars have used these differences to classify people into various races. They have devised racial categories for human beings according to such physical characteristics as the color of the skin, the color and texture of the hair, and the shape of the eyes.
But some people assigned to the same race—and even some members of the same family—have widely differing features. Over the years, scientists have disagreed over how many races of human beings can be devised, and over which individuals belong to what race. For this reason, many anthropologists and biologists have come to believe that the assignment of a racial label to any group of people is arbitrary and thus open to argument.
For many years, most scholars believed that “pure” races of human beings existed some time in the prehistoric past. According to these scholars, the “pure” human races developed in complete isolation from one another, and the members of each race exhibited physical characteristics that the members of other races did not possess.
Today, however, most physical anthropologists (scientists who study the physical differences and prehistoric development of human beings) doubt that “pure” races ever existed. They point out that people have probably always taken mates from outside their own population as well as from within. They also note that as transportation and communication have become easier, populations have blended more and more. For these reasons, the biological definition of race does not describe human populations well. Most anthropologists now avoid classifying people into races. Instead, they try to learn more about human diversity by studying how human traits vary throughout the world.
Despite the lack of a scientifically valid racial classification system, people generally consider those who “look different” from themselves to be members of a different race. As a result, the concept of race remains important in a sociological sense. Societies continue to divide their members into “races”—though the criteria and labels used may vary from society to society.
The idea of race has often been misunderstood, and the term has sometimes been misused on purpose. The biological concept of race has often been confused with culture, language, nationality, or religion. Differences in physical appearance have led some people to mistakenly conclude that members of different groups are born with differences in intelligence, talents, and moral standards. Race has also been a major basis of discrimination—that is, the treatment of other groups as inferior to one’s own group. For more information, see the World Book articles on Minority group, Racism, and Segregation.
Systems of racial classification
Physical differences among human beings have long been recognized, and many of these differences have been used throughout history as bases of racial classification. Obvious physical characteristics, such as size, build, skin color, eye form, hair form, and nose shape, were the main criteria of early classifications of race, with skin color considered most important.
Since the beginning of recorded history, scholars have classified human beings in different ways, and the number of categories recognized by each system varied. The development of racial classification systems was influenced by three important theories: (1) the three-race theory, (2) evolutionary theory, and (3) the geographical-race theory.
The three-race theory.
Ancient Egyptians, Greeks, and Romans knew about dark-skinned, curly-haired peoples that lived in Africa. They also knew about the so-called “yellowish-skinned” peoples of Asia, most of whom had folds of skin that extended from their eyelids over the inner corners of their eyes. Limited knowledge of the peoples of the world at this time suggested the existence of three races—European, or “white”; African, or “black”; and Asian, or “yellow.” These groups eventually became known as Caucasoid, Negroid, and Mongoloid, respectively. For many years, scholars attempted to classify all human populations in terms of these three races, or some variation of the three. They believed that all people belonged to one of a limited number of racial types. They also believed that the traits of each race were fixed and unchanging.
The major period of European overseas exploration, which began in the late 1400’s, provided increased contacts with peoples of different cultures. By the 1800’s, it became evident that much of the world’s population did not easily fit into the three-race system. For example, as Europeans came into contact with more and more Asian peoples, they realized that the skin of the people they had classified as Mongoloids was not really yellow, but that it varied from very dark to very light brown. They also discovered that the epicanthic fold—the inner eyefold thought to characterize Mongoloids—was rare in some Asian populations but present in some of the native peoples of southern Africa and North America. Lip form and hair form were also found to vary across the traditional racial groupings.
Evolutionary theory.
The view that human beings could be classified into races based on fixed physical characteristics began to change dramatically as biologists came to accept the theory of evolution. During the 1700’s, most biologists believed that all plant and animal species remained the same from generation to generation. However, geologists of the early 1800’s found fossils of animals and plants that were not the same as living species, thus providing evidence that species were not fixed.
Even though scientists could now see that species could change, they did not know how evolution worked. It was the idea of natural selection as the mechanism for evolution that helped scientists understand how organisms could change over many generations. This idea, set forth by the British naturalist Charles R. Darwin in his book The Origin of Species (1859), states that populations of organisms can change over generations as they adapt to their physical environment. This new understanding of the processes of evolution through natural selection, when applied to human populations, showed that many of the supposedly “fixed” traits that had been used to identify races were actually adaptations that had evolved over time in response to environmental conditions. See Evolution; Darwin, Charles R.
Scientists saw that widely separated groups could develop similar characteristics as a result of adapting to similar environments, even if they shared no recent ancestral relationship. For example, the Quechua, a people who live in the Andes Mountains of South America, and the Sherpas, a people of the Himalaya in Asia, are only remotely related. However, they have many similar physical characteristics as a result of prolonged adaptation to living in their high mountain environments.
As they came to understand evolutionary theory, experts began to see the difficulty of trying to use adaptable traits to fit people into just a few major races. Physical anthropologists began to search for nonadaptive, or neutral, traits—that is, physical characteristics that would persist even if a population moved to a different environment. They viewed race as something fixed and unchanging and wanted to discover traits that were also unchanging. Anthropologists compared many traits and physiological processes of people living in different environments, including blood groups and rates of respiration, circulation, and metabolism. These comparisons are discussed later in this article, in the section on How human populations develop and change.
The geographical-race theory.
In an effort to reconcile the theory of evolution with the observed variations among the world’s populations, some anthropologists developed a new system of racial classification during the 1950’s. They divided human beings into large categories called geographical races. These races were collections of populations that exhibited similar characteristics. One popular classification system recognized nine geographical races: (1) African, (2) American Indian, (3) Asian, (4) Australian, (5) European, (6) Indian, (7) Melanesian, (8) Micronesian, and (9) Polynesian.
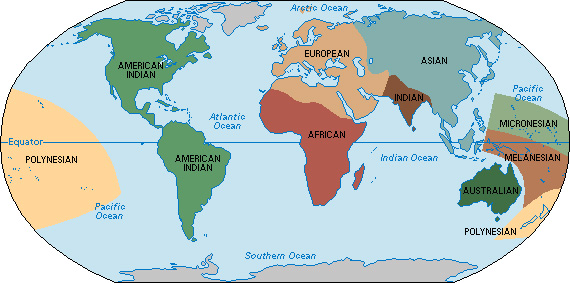
In general, the geographical races extended throughout major continental areas and large island chains. But they did not correspond exactly to the continents. For example, the European geographical race included populations throughout Europe, in the Middle East, and north of the Sahara in Africa. It also included descendants of these populations in other parts of the world, such as the “whites” of North America and Australia.
Geographical races were believed to exist because of the isolation caused by such natural barriers as oceans, mountains, and deserts. The idea was that these barriers separated groups of people for many thousands of years, allowing the populations to evolve in different directions. India, for example, is partly isolated from the rest of Asia by the Himalaya. According to the geographical-race theory, this isolation permitted the Indian geographical race to develop separately from the Asian geographical race.
Anthropologists used the term local races to describe distinct subcategories of geographical races. Some local races had millions of members. For example, the Northwest European local race included the populations of Scandinavia, Germany, Belgium, Luxembourg, the Netherlands, Great Britain, and Ireland. It also included peoples who emigrated—or whose ancestors emigrated—from those areas. Local races containing much smaller numbers of people included the Sami (formerly known, to outsiders, as Lapps) of extreme northern Europe and the Basques, who live in northern Spain and southwestern France.
Some anthropologists used the term microraces to describe the subpopulations that existed within local races. But microraces—and even local races—could not always be clearly defined. Within a given geographic area, the members of different subpopulations often intermarried, so the physical features used to define these groups blended together.
This expanded, detailed classification system represented a major change in the view of human races. The geographical-race system took into account the theory of evolution as well as heredity, recognizing that populations are shaped by their environment. However, many anthropologists believed it did not eliminate the problems of the older systems. Because members of different races could possess the same physical characteristics, the racial criteria could not be clearly identified.
Alternatives to racial classification
In the past, scholars based racial classifications on clusters of physical characteristics that supposedly represented the “typical” member of that race. But many of the individuals categorized in a particular race did not reflect all the characteristics attributed to that race. In addition, the scholars who constructed classification systems did not always agree on which traits—or how many—should be considered.
To see the problems involved in defining races by means of “typical” characteristics, consider skin color. A pigment called melanin determines skin color. Dark skin contains more melanin than light skin does. Skin color has been used as a major classifying characteristic in all racial systems. For example, a light brown skin color was considered “typical” for the members of the European geographical race. But some members of the race had skin that was far lighter than the “typical” color, and others had skin that was much darker. Similarly, the members of the African geographical race “typically” had brownish-black skin. But again, many individuals classified in this race had skin that was lighter than the “typical” shade, and many others had skin that was darker.
To further confuse matters, some of the darker-skinned members of the European geographical race had skin as dark as some of the lighter-skinned members of the African geographical race. In view of these complications, it has become extremely difficult to assign people to a race based solely on skin color.
Increasing the number of identifying traits only added more problems. The shape and fullness of the lips, for example, varied widely among people who were considered members of the same race. Furthermore, lip shape demonstrated the same kind of overlap among members of supposedly different races as did skin color.
These problems have led many anthropologists to conclude that classification based on physical characteristics is not scientifically valid and serves no useful purpose. They find the study of human variation to be more productive than the assignment of racial labels. As a result, they have adopted alternate approaches to traditional systems of racial classification. Chief among these alternatives are (1) the clinal approach and (2) the population approach.
The clinal approach.
The geographical distribution of a physical characteristic can be shown on a map by zones called clines. Clines are formed by drawing lines to connect points of the same or similar frequency. For example, in the case of skin color, each cline includes locations at which populations demonstrate the same average skin color. As variations from dark to light are plotted on a map, certain distribution patterns begin to emerge. A clinal distribution does not associate specific traits with traditional racial categories, nor does it associate different traits with one another. For example, skin color and blood type would be plotted on separate maps and show different patterns of distribution.
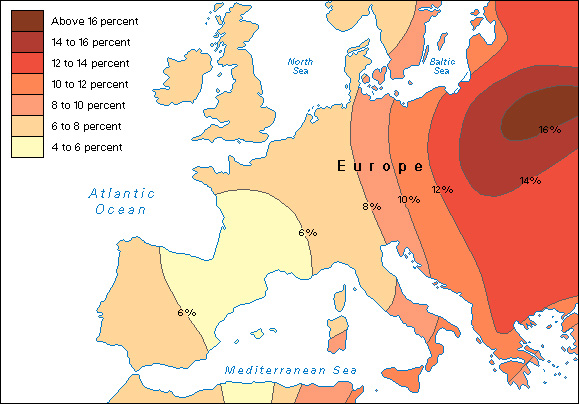
The clinal approach has been used extensively to examine the worldwide distribution of blood types. Scientists classify human blood into groups according to proteins on the membranes of the red blood cells. The presence or absence of these proteins is determined by heredity. Studies show differences in the frequencies of some blood groups throughout the world.
The best-known blood-group system is called the ABO system. In this blood-group system, type O is the most common. Type O is followed by types A, B, and AB. Other systems that are used in comparing blood-group frequencies include the Kell, Kidd, Lutheran, MNS, P, and Rh systems. See Blood (Blood groups).
Clines of blood-type distribution help anthropologists consider possible explanations for the geographic variations they observe. For instance, clinal mapping shows that central Asia has the lowest frequencies of type O blood. One possible explanation for this has to do with the deadly epidemic disease bubonic plague—a disease that has long been present in central Asia. The surface proteins that characterize type O red blood cells resemble the surface proteins found on the infectious bacteria that cause bubonic plague. Normally, the body can produce disease-fighting chemicals that recognize and attack cells that carry the bubonic-plague surface proteins. But if a person has type O blood, the body is less likely to make these disease-fighting substances because they would damage its own red blood cells. During a plague epidemic, central Asians with type O blood would have been at greater risk of dying from the disease than were those with other blood types. Over the centuries, this disadvantage could have led to the comparatively low frequency of type O blood in central Asia.
The population approach
is used to study patterns of variation among human populations. Anthropologists define a population as a group of similar people who are more likely to mate with one another than with outsiders. Anthropologists using the population approach investigate clusters of physical traits but make no assumptions about race on the basis of those clusters. Instead, they see each population as the product of a unique set of circumstances, including adaptation, genetic change, isolation, and history of migration. These researchers then attempt to explain the similarities and differences among the populations. They do not try to fit the populations into racial categories.
The population approach assumes that groups of people who have lived in similar environments for a long period will demonstrate similar adaptations. This can happen even if the location of these similar environments is far apart. For example, populations living at very high altitudes must adjust to extreme conditions. Temperatures can get extremely hot during the day and very cold at night. Also, the air pressure is so low that less oxygen is available, making breathing more difficult. Throughout the world, populations living at high altitudes show specific traits in response to similar environmental conditions. For instance, their lungs can hold more air than those of people at lower altitudes, enabling them to inhale more oxygen with each breath.
How human populations develop and change
The characteristics studied by physical anthropologists—such as eye color, nose shape, blood type, body height, and susceptibility to genetic diseases—are determined by both heredity and the environment. The inherited aspects of a trait are determined by tiny biochemical structures in cells, called genes. Genes contain chemical instructions for the formation of hereditary characteristics. Children inherit half their genes from their father and half from their mother. The underlying genetic makeup of a trait is called the genotype. The actual appearance of the trait is called the phenotype. The phenotype results from the environment and heredity.
Members of the same population of human beings tend to have more genes in common than do members of different populations. Closely related populations also share more genes than do distantly related groups, just as cousins have more genes in common than do members of different families. All the genes in a population are called the group’s gene pool. The degree to which a gene is present in a population is called the gene frequency. For more information on how characteristics are inherited, see Heredity; Cell; Gene.
Scientists have shown that the gene pools of human populations can change over time. The presence of some genes increases, while the presence of other genes declines. As gene frequencies change, the frequencies of physical characteristics in a population may also change. Such changes can result from a number of different factors, including (1) natural selection, (2) mutation, (3) genetic drift, (4) the founder effect, and (5) migration and gene flow.
Natural selection
is the process that enables some organisms or individuals to live and reproduce while others do not survive. Those who reproduce pass their genetic characteristics on to their offspring. Natural selection is the force that drives Darwinian evolution. For example, certain individuals within a population might possess a genetic characteristic that provides resistance to a local disease. As a result, those individuals tend to survive longer and to produce more offspring than the other members of the population. Moreover, their children who inherit the favorable characteristic will likewise tend to live longer and leave more descendants. Over time, individuals who possess the favorable trait will tend to outnumber those who do not, and the gene frequencies of the population will have changed.
As a result of natural selection, a population that lives in a certain area for many generations tends to exhibit distinctive genetic traits or clusters of traits. Scientists have shown that differences in skin color, body build, and other physical characteristics represent adaptations to environmental factors. See Natural selection.
Climatic adaptations.
The genetic makeup of populations may change over time to adjust to climate. For example, dark and light skin and eye color represent adaptations to different amounts of sunlight. The color of our skin, hair, and eyes is determined by the pigment melanin. The amount of melanin in the skin, hair, and eyes can differ greatly from one person to another. Large amounts of melanin in the skin help protect it from sunburn and reduce the risk of skin cancer. Dark pigment in the eyes improves vision in bright sunlight. Therefore, dark skin and dark eyes represent adaptations of people whose ancestors have lived for many generations in sunny climates.
Sunlight also affects skin color in another way. Our bodies need vitamin D to help us absorb calcium. The absorption of sunlight enables our bodies to make vitamin D. In climates with long winter nights, it can be difficult for our bodies to absorb the sunlight needed to make enough vitamin D. People whose ancestors have lived in these climates for generations have adapted to reduced sunlight by developing light-colored skin that will absorb the little sunlight that is available. Therefore, skin colors in humans result from adaptations to the environments in which our ancestors have lived.
Human populations also differ in response to cold. Among Inuit, for instance, the body maintains a high temperature by burning large amounts of fat and protein. (Inuit are the people formerly called Eskimos by outsiders.) It also keeps large amounts of blood flowing to the arms, legs, fingers, and toes, to prevent frostbite. Aboriginal Australians, who live in a generally warm climate but traditionally slept in below-freezing temperatures with little clothing or shelter, have adapted to cold in a different way. The temperature in their legs and feet drops during sleep, and they burn less energy. But their bodies maintain warmth in the trunk, where the vital organs are.
The Aboriginal peoples of Australia and the Inuit of the Arctic have both adapted to cold. The Aboriginal peoples had a limited food supply. They had no extra food to burn for body heat when the temperature dropped. Instead, their adaptation enables them to save body energy. But this method would not work in the Arctic, where the climate is cold day and night. The Inuit are adapted to extreme cold—temperatures as low as -40 °F to -60 °F (-40 °C to -51 °C). Their adaptation depends on the availability of food to supply energy and body heat.
Susceptibility to genetic diseases.
Many of the diseases that afflict human beings have some genetic basis. Human populations differ in the frequency of the genes that cause certain genetic diseases and disorders. For this reason, a number of genetic diseases are distributed differently throughout the world and affect some populations more than they do others. The fact that certain populations are plagued by particular genetic diseases can be explained in terms of natural selection.
The frequency of a hereditary blood disorder called sickle cell anemia varies widely in different populations. Individuals who inherit the sickling gene from both parents suffer from sickle cell anemia. Most cases of this disease are fatal. Carriers—people who inherit the defective gene from only one parent—may have almost no problems or experience only mild symptoms. But they can transmit the abnormal gene to their children. See Sickle cell anemia.
Scientists have found that carriers of the sickling gene have a higher resistance than noncarriers to malaria, a dangerous disease transmitted by certain mosquitoes. Sickle cell anemia is a rare disorder, but it occurs more often among populations of western Africa, the Middle East, southern Europe, and the Caribbean, most of whom live in areas threatened by malaria. Thus, the sickling gene—despite its negative effects—represents an important advantage for people in these areas.
Loading the player...Malaria growth
Another genetic disease, cystic fibrosis, is more common among European populations and their descendants than among other populations. This rare and incurable disease affects the lungs and other organs. Like sickle cell anemia, cystic fibrosis results from the inheritance of a disease-causing gene from both parents. Carriers of the gene do not contract cystic fibrosis, but they can pass it on to their offspring. Some scientists believe that carriers are more resistant than other individuals to tuberculosis, an infectious disease affecting the lungs. Tuberculosis swept through Europe from the 1700’s to the early 1900’s. Carriers of the cystic fibrosis gene may have been more likely to survive these epidemics, which would explain the relatively high frequency of the cystic fibrosis gene in these populations. See Cystic fibrosis.
Mutation.
A mutation is a change in genetic material. A changed gene often produces a different inherited trait that can be passed on to future generations. Mutations result from a chemical change in DNA (deoxyribonucleic acid), the chief chemical compound of genes. Mutations may also result from a change in the number or arrangement of chromosomes, the threadlike structures that contain the genes. Scientists know of many agents that can cause mutations, such as certain types of radiation, chemical treatments, and heat, but they cannot tell in advance which genes or chromosomes will mutate or how the trait controlled by that gene or chromosome will change.
Many mutations are harmful, causing mental or physical disorders. But other mutations are neutral, and some are favorable. A favorable mutation may provide the raw material for natural selection by making a person better suited to the environment. For example, a mutation that enhances the body’s ability to make vitamin D from sunlight would be advantageous to a person living in the Far North, where the earth receives less sunlight. Such beneficial genes will increase in frequency from generation to generation. On the other hand, individuals possessing harmful mutations may be selected against, so the trait will not tend to increase in a population. In this way, mutation sometimes works together with natural selection to produce changes in gene frequencies. See DNA; Heredity (Sources of genetic variation); Mutation; RNA.
Genetic drift
refers to chance fluctuations in the gene frequencies of a population from generation to generation. The genes of each generation represent only a sample of the previous generation’s gene pool. As a result, the gene frequencies of each generation of individuals tend to vary randomly within the limitations of the preceding generation’s gene pool. The smaller the population, the stronger the impact of these fluctuations is likely to be. Such changes are not likely to have much effect in very large populations, but they can lead to significant genetic changes in small populations.
The founder effect.
When a small number of people from a large population establish a new population in a different place, it is unlikely that the founders represent the full range of diversity in their parent population’s gene pool. When these founders produce offspring, a smaller, more limited gene pool is created. This phenomenon is called the founder effect. In future generations, the members of a population influenced by the founder effect are likely to resemble one another more closely than they do the members of the larger, more diverse parent population.
The founder effect may explain the increased incidence of certain traits or diseases in a population. For example, a hereditary brain disorder called Tay-Sachs disease occurs mainly among Jewish children of eastern European ancestry. People with one Tay-Sachs gene do not have the disease but may transmit the gene to their children. Children who inherit the gene from both parents have the disease. The Jews of eastern Europe made up a small population with a limited gene pool, so the incidence of the disease remains higher among their descendants than in other populations. See Tay-Sachs disease.
A similar limitation on a population’s gene pool may occur if the genes of one person or family in a small population are passed to a large number of offspring. For example, if one man in a small, isolated group married several women and fathered many children, his genes would appear in future generations with more frequency than would the genes of other members of the population.
Migration and gene flow.
When migration occurs between separate populations, new genes or combinations of genes are likely to be introduced into each group through interbreeding. As a result, the gene pool of each group comes to include genes from the gene pools of the other populations. In this manner, migration may cause the gene frequencies of populations to change over time. In modern times, easy access to transportation has greatly increased gene flow.
Since earliest times, people have moved from one place to another and have chosen mates from other groups. The greatest amount of gene flow occurs between populations that live next to one another. Mixture may also occur as a result of various cultural practices. Throughout history, such practices as exploration, colonization, bride capture, and enslavement have brought individuals of various genetic makeups together. The result in many cases has been change in the gene frequencies of the populations affected by these practices.
The social significance of race
As we have seen, most physical anthropologists have abandoned the idea of classifying human beings into biological races. In many societies, however, people continue to identify themselves and others as members of a particular race, often based on skin color. Thus, whatever its shortcomings on a biological basis, racial classification remains an important sociological factor. Social scientists must recognize the way a society defines racial categories if they hope to understand human behavior. It would, for instance, be difficult to analyze American society without taking into account the commonly used division of that society into “white,” “Black,” “Hispanic,” and other races. Yet these categories themselves reveal problems with the concept of race. “White” and “Black” represent categories traditionally used to identify biological races. But “Hispanic” refers to the language group of Spanish-speaking people, not to any one biological group. Unfortunately, many social distinctions between races result from racial prejudice and misunderstanding.
Race and ethnic or national identity.
The biological concept of race is sometimes confused with the idea of ethnicity or nationality. People identify themselves as members of certain ethnic or national groups based on certain geographical, cultural, or religious characteristics. However, these identifications are not based on physical differences. For instance, people sometimes incorrectly speak of the “Arab race,” the “German race,” the “Irish race,” or the “Jewish race.” But these labels refer to ethnicity or nationality and have nothing to do with the biological concept of race.
Race and discrimination.
History includes many episodes in which the members of one group of people deemed themselves superior to another group. Such beliefs were long used to rationalize the enslavement and persecution of people viewed as inferior. For example, the ancient Romans viewed the Germanic tribes as a “race” of barbarians—that is, non-Romans—who were barely human. Europeans who settled in America claimed superiority over the Indigenous (native) people to justify their expansion into the New World. In the 1930’s, the leaders of Nazi Germany preached that Germans belonged to the “superior Aryan race,” and that Jews and all other non-Aryan peoples were inferior.
Experts have not discovered any scientific basis for such claims of superiority. But many people still view other groups in terms of stereotypes. That is, they have oversimplified, preconceived, and generalized beliefs about the members of these groups. At various times, for example, certain groups have been described as dirty, dishonest, sly, humorless, or dull. These judgments have often been confused with racial traits, though they have nothing to do with the biological concept of race. Many such judgments have nothing to do with culture either, but only with the opinions or prejudices of those who make them. Discrimination can result from these stereotypes. As a result of these beliefs, members of minority groups in many societies have fewer educational and job opportunities than do members of the majority group.
The belief that some groups are more intelligent than others has been used to justify discrimination. Scientists have shown that a person’s intelligence is partly inherited and partly determined by the environment. The use of intelligence to compare groups of people is extremely difficult, because few such comparisons can be considered equal. A better-educated group, for example, will score higher on tests that measure education. Groups that value mathematical skills or technical ability will do better on tests involving such skills.
Many experts believe it is impossible to design an intelligence test that is not influenced by a person’s experiences. Nevertheless, scientists are trying to develop culture-fair or culture-free tests that reduce the effects of cultural differences on test scores.
The differences among human beings make the world a fascinating place in which to live. But when people focus on these differences, they often fail to appreciate how similar all human beings are. Most of the distinctions people make between themselves and others have much more to do with culture than with biology.
